

Reproductive habits and first feeding of the marine ornamental, Orchid Dottyback (Pseudochromis fridmani)
Master's Thesis by Josh Borland
Supervisors: Chaoshu Zeng, Bill Chen
Master's Thesis by Josh Borland
Supervisors: Chaoshu Zeng, Bill Chen
Abstract
Marine ornamental aquaculture is a young and growing field in which efforts to captively rear and breed fish are being investigated. These efforts have both economic and ecological significance as they provide alternatives to wild harvest which can devastate reef environments. Part of this effort is the continued research into improving culturing techniques for the targeted ornamental fish. Previous studies have identified the first feeding of larvae and broodstock establishment as the primary bottlenecks in production. This study observed 13 pairs of Pseudochromis fridmani broodstock to determine their reproductive habits and outline their commercial viability. It was determined that once established, P. fridmani can be expected to spawn every 7-11 days and each spawn then has ~85% of lasting until hatching. However, establishing a pair can be difficult to do quickly as they lack any obvious sexual dimorphism and even when paired appropriately take a while before they can maintain a clutch successfully to hatch. This study also tested six different feeding treatments (Rotifers at 2/ml, 20/ml, and 40/ml; and Copepods at 0.1/ml, 0.5/ml and 1/ml) to determine the optimal prey density for P. frimani’s first feed. The mean number of prey within the gut was used as a measure of feeding success. It indicated that rotifers at a density of 20/ml was the best density and 1/ml was the best for copepods. This suggested some form of feeding inhibition occurs at high rotifer density, because otherwise gut content would be expected to be the same or greater than that of 20/ml. For copepods, increasing the density further would likely continue to be matched by increased feeding, but these densities are likely unrealistic given the difficulties associated with copepod culturing. Finally, additional experiments were proposed. One to improve broodstock establishment with light and temperature manipulations and another to test first feeding response with mixed prey options to test for prey preference or increased feeding success.
Introduction
Marine ornamental aquaculture (MOA) can be broadly defined as: the effort to captively rear and breed organisms (predominantly fish) that are bought and sold within the Aquarium trade. MOA is a relatively young field of growing importance as demand for ornamental fish continues to rise (Ostrowski & Laidley 2001). Studies show that unregulated collection from the wild continues to be a problem as the methods used are often destructive to the reef environment and unsustainable in nature (Abraham et al. 2017; Moorhead 2015; Ostrowski & Laidley 2001). Currently an estimated 10% of marine fish within the trade are sourced from captive production, despite the fact that the MOA fish generally fare better and survive longer in captivity, and experience reduced shipment stress (Olivotto et al. 2011). Studies also find wild sourced fish tend to have high mortality rates from capture to the home aquarium, in some instances up to 73%; but captive breeding and domestication within MOA would likely reduce this mortality rate (Stevens et al. 2017). These are some of the reasons MOA is particularly important as it offers a practical alternative to wild harvest, while providing other benefits.
MOA not only has ecological value, but a large economic potential as well. Currently MOA is valued at nearly 15 billion USD and is growing steadily (Moorhead 2015). This is primarily due to the large economic value of the fish being traded. A marine ornamental’s worth, on average, is $500/kg compared to that of food fish at $6/kg (Morcom et al. 2018). MOA also represents potential savings in regard to product loss during handling and delivery. The high value of marine ornamentals and the high mortality of wild sourced fish represent a monumental loss of potential income which could be reduced through the substitution of captively bred fish which tend to be more durable and stress tolerant (Stevens et al. 2017).
Unfortunately, despite its ecological and economic value, very little research exists in regards to most of the marine species in the aquarium trade. Studies of the few species that are better understood have revealed that the primary limitation of production is the first feeding of recently hatched larvae (Abraham et al. 2017; Olivotto et al. 2011; Ostrowski & Laidley 2001). This difficulty of culturing the early life stages is primarily a function of the larvae’s extremely small mouth gape and need for live feeds (Abraham et al. 2017). Some studies have provided much needed insight into this field, particularly studies looking at relevant prey items for such small fish. Rotifers, copepods and Artemia are naturally occurring prey which have become commonly used.
Artemia, while commercially the easiest to provide are quite large and usually unsuitable for first feeds (Sorgeloos et al. 2001). Conversely, rotifers and copepods are significantly smaller and therefore, predominantly used due to their small size. In relation to rotifers, while not nutritionally the best, they are easy to culture at high densities (2000-20,000/ml) and could easily be provided at densities higher than naturally occurring to encourage feeding and determine the likely optimal feeding density (Hagiwara et al. 2001). Copepods on the other hand, while much more nutritious, are significantly more difficult to maintain in high densities, at which egg cannibalism becomes a serious problem. Furthermore, when compared to rotifer densities capable of reaching concentrations of >2000/ml, the “high density” copepod cultures of ~3.5/ml are very low, so the testing densities must be similarly low to be commercially relevant (Vu et al. 2017). Despite the wealth of knowledge on culturing these prey items, knowledge concerning optimal feeding density for larvae is largely unavailable.
Further complicating the situation, other culturing difficulties can occur in the adult stages of the fish, such as difficulty sexing mature fish, or meeting the nutritional needs of broodstock with commercially available feeds (Olivotto et al. 2011). To be commercially viable, a species ideally needs to: provide larvae regularly or on demand, have a financially sustainable survival rate, and demand within the aquarium trade. Assuming a fish is in high demand, understanding its reproductive habits can elucidate the general availability of larvae; if determined to be low, can this be modified with intervention such as induced spawning either chemically or with environmental cues such as simulated seasons (Hill et al. 2009). These issues provide a clear knowledge gap and outline the general need for studies in this area.
Within these areas of broodstock establishment and larval first feeds, there has already been some progress made in regards to select species. This study will focus on the Orchid Dottyback (Pseudochromis fridmani, or P. fridmani), a marine fish from the Red Sea. Previous studies have noted that this species is protogynous hermaphroditic and capable of bi-directional sex change allowing for pairing despite the initial sex of the pair (Mies et al. 2014). While P. fridmani has been successfully cultured previously, cannibalism of eggs and poor larval success has prevented it from becoming successfully cultured worldwide (Mies et al. 2014). This study is part of an effort to fill these knowledge gaps.
In this study the general reproductive habits of P. fridmani will be observed and then an experiment looking at the first feed will be conducted to see how it might be optimized. Different concentrations of two commercially available and relevant prey items (rotifers and copepods) will be provided to test for differences exhibited in the first feeding by P. fridmani larvae. When investigating these feeding densities, it is expected that the feeding protocol for larvae might be optimized allowing for a more stable culturing effort. By providing the appropriate density of feed for larvae, it can be expected that they will have reduced early mortality thereby, increasing the chances of surviving to a commercially viable age. Finally by tracking the broodstocks reproductive habits, it is expected that the average spawning interval, hatching success, and reproductive potential will be discovered. This will allow for commercial providers to make educated decisions when beginning cultures of P. fridmani and inform on optimal feeding density once a stable larval production is established.
Methods
Broodstock maintenance and observation
Broodstock were organized into thirteen pairs. Some pairs were pre-existing from previous studies and were not separated to minimize interruptions in reproduction. One such pair included a third fish, but due to the limited differences in sexual dimorphism it was impossible to isolate the duplicate sex and remove it, so was left together as a triplet. Due to limited space and available research tanks, ten pairs were isolated in replicate conical tanks of 120L and the three other pairs (including the triplet) were kept in a different set of replicate tanks, which were much larger (2000L) and differently shaped (rectangular) (Fig. 1). All the tanks were outside with water temperature and quality maintained by the Marine Aquaculture Research Facility Units (MARFU) recirculatory system. Ideally, this meant the conditions stayed between 27.5 to 29.5 °C, 29 to 37‰ salinity, 8.0 to 8.2 pH, NH3and NO2 <0.02 ppm, and NO3 <6 ppm. All illumination was the result of a natural photoperiod.
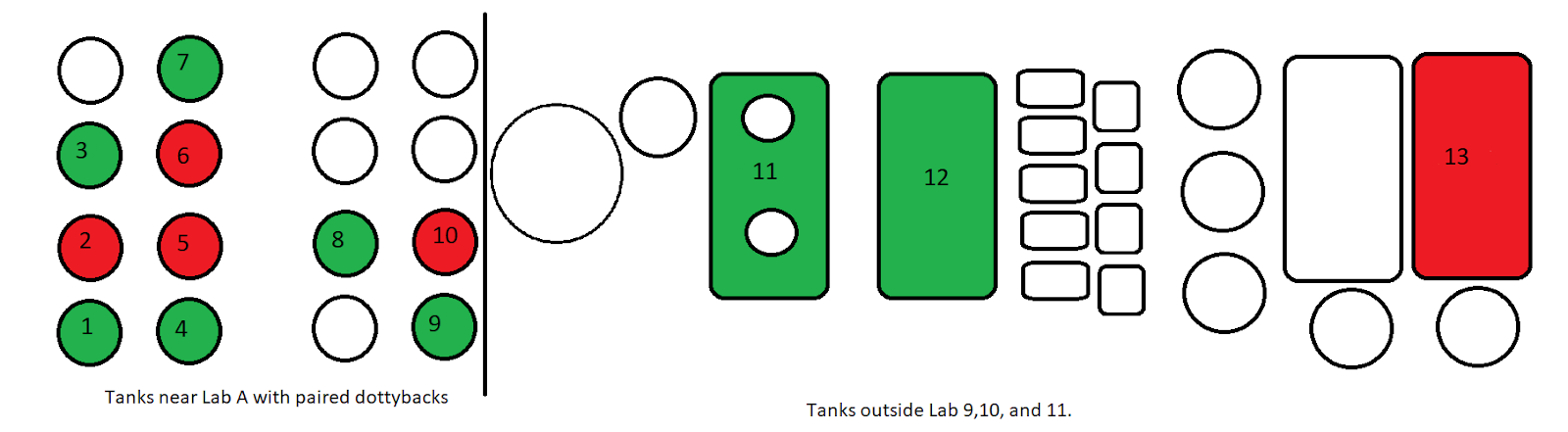
|
| Figure 1. Diagram of tank layout within the MARFU complex. Green tank housed pairs that successfully spawned. Red tanks never spawned. Note red tanks seem isolated within center of Lab A complex. |
Within each tank, the broodstock were provided a PVC “den” in which the male tends to spend the majority of its time. These pipes were approximately 25cm long and 40mm in diameter. On one end was a cap to provide a fully enclosed area and the other end had a reducer. Importantly this “entrance” was smaller in diameter (25mm) than the interior to provide a sense of security to the adult fish (Fig. 2). Also within each tank was a bundle of seagrass to provide shelter to the female and habitat complexity. Other fish were also present in the majority of the tanks, but no competing pairs of demersal spawners were present. To monitor the reproductive behavior, the pipes were visually checked for eggs twice a day; once in the morning between 8:30-10:30am, and once in the afternoon between 3-5pm. This data was collected for each of the thirteen pairs for 90 days beginning July 22nd through Oct 20th 2018. During this time fish were provided with a broodstock feed, developed by Moorhead & Zeng 2011, twice a day. Data was analyzed with Microsoft Excel 2013.

| 
|
| Figure 2. Photo of the PVC dens provided to the P. fridmani broodstock. The exploded view provides a good view of the difference in entrance and internal diameters. |
In an effort to record the spawning behavior, one pair was also filmed. This was achieved through the use of a GoPro Hero 3+ hung near the surface of the tank facing down at the PVC den. Unsure of when the fish would spawn the camera started recording at noon the day anticipated the fish would spawn given their previous spawning had been 7 days prior. To ensure the behavior was captured the camera was left recording, and the battery swapped every hour, until the afternoon check at 5 pm when it was confirmed that the fish had spawned. The footage was later reviewed using GoPro video editing software.
Larvae-First feed
To first establish if the methods described by Hilder et al. 2014 would be applicable to P. fridmani, a series of pilot trials was conducted. One concern was whether or not this species would eat enough of the prey items provided within four hours to allow for comparison. Another concern was whether or not the euthanization protocol would cause the larvae to spill their gut prior to analysis. The final concern was whether or not the different prey items would be identifiable post-ingestion. To ease these concerns multiple small scale pilot trials were conducted. The first tested and confirmed that the euthinization protocol would work and that identification of rotifers was indeed possible post ingestion. The second trial revealed that differences in gut content were present after only four hours between treatments. The final treatment was perhaps the most important as it revealed that cofeeding caused difficulties in identifying if the prey item within the gut was a copepod or a rotifer. This was particularly difficult with the different larval stages of copepods which were closer in size to rotifers than an adult, which were more easily distinguished. Given this result, cofeeding treatments to distinguish if larvae have a prey preference were excluded. Having shown the various treatments were likely to be informative, the full experiment could now be conducted.
The larvae used in the first feed studies were acquired from the broodstock previously described. On the 4th day post fertilization (DPF), the PVC “den” with the egg bundle and male were removed from the tank and introduced to a hatching tank (25L) around sunset (5-7pm). This hatching tank was completely black and then covered to eliminate any light and left for a 3-4 hours to allow hatching (Fig. 3). Approximately an hour after hatching (roughly 8pm), the cover was removed and the PVC den with the male and any unhatched eggs were removed from the tank. Light aeration was provided overnight.

|
| Figure 3. Photo of the hatching tank with cover in place. |
The next morning, 18 experimental tanks were prepared, to allow for three replicate tanks for each of the six feeding treatments. Each tank had a final volume of 3L and included a mix of algae paste (RotiGrow Nannochloropsis sp., ~5.0X106cells/ml) and live algae (Ishochrysis sp., ~5.0X105cells/ml) to green the water (total density 5.3-5.5X106cells/ml), and dark plastic exterior to reduce photo stress (Fig. 4). Then 25 larvae were randomly allocated to each tank, which were randomly assigned to one of six treatments. Because of the limited larvae availability, it was impossible to reach the desired density of 10 larvae/L which has been commonly used in previous first feeding experiments (Degidio et al 2017; Hilder et al. 2016). To ensure no tank had a longer acclimation period larvae were randomly assorted between tanks as the transfer process was quite lengthy. All the larvae were distributed to experimental tanks by 10:30 AM September 12, 2018. After a 30min acclimation period, the prey was introduced with 5 minute gaps between treatments. The rotifer densities were 2/ml, 20/ml, and 40/ml; while the copepod densities were 0.1/ml, 0.5/ml, and 1.0/ml. Note that mixed culture treatments were considered, but during the trial phase it was determined differentiation of rotifers and copepod nauplii post ingestion is quite difficult, so to prevent confounding errors, cultures were kept separate. The prey densities were determined primarily based on the relevance to the commercial facilities as previously discussed. Then after 4 hours (in the same order) the larvae were euthanized with Aqui-S according to manufacturing instructions to avoid regurgitation of prey from more abrupt euthanasia methods (Degidio et al. 2017). During the feeding period, there was no water exchange, or change in lighting conditions which might affect feeding. After euthenization, the larvae were transferred to formalin and stored at 2 °C to wait analysis.

|
| Figure 4. Photo of final experimental setup. Each 3L tank was individually wrapped in dark plastic to provide a consistent light level. To distinguish between tanks and treatments, yellow labeling tape was used. Josh is recording the current time as the feeding was just initiated. |
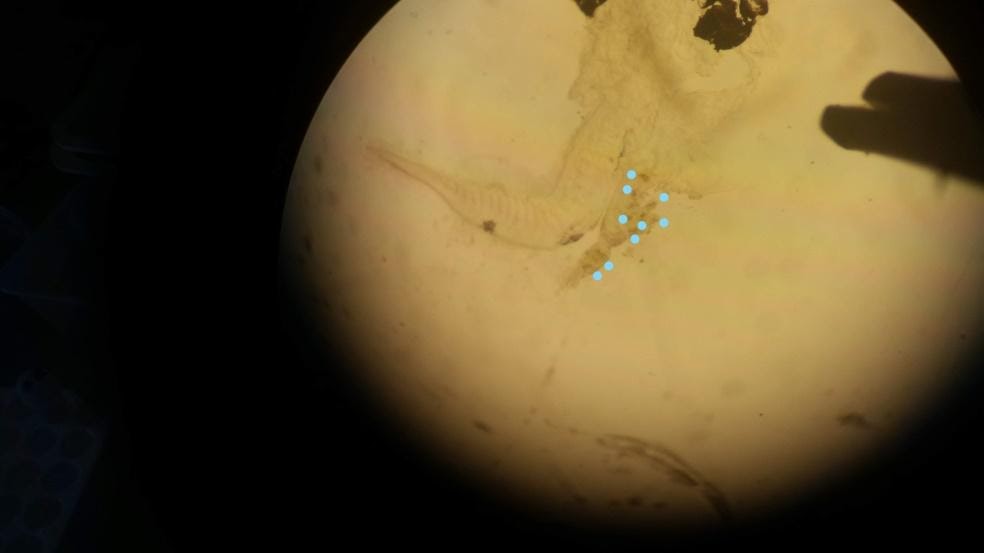
To analyze the gut content, the larvae were “squished” between a slide and cover slip rupturing the gut and exposing its contents and then the number of rotifers or copepods within were counted under magnification (Fig. 5) (Degidio et al. 2017). The data was tabulated and analyzed via IBM SPSS 2017. To determine if there were significant differences between gut content within treatments, ANOVA was used. Additionally, the percent of larvae that had an empty gut were also compared using ANOVA. For each statistical test all assumptions of randomness, normality and heterogeneity were met unless otherwise noted.
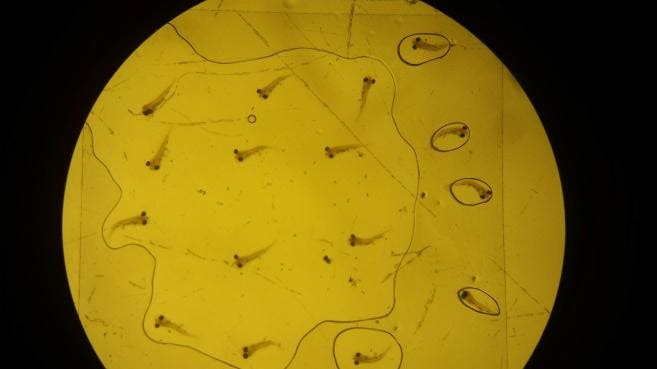
| 
|
| Figure 5. A) Photo of trial larvae under magnification. For experimental samples the magnification is higher and forceps are used to “squish” the larvae until the rotifers or copepods are more apparent and quantifiable. B) Experimental larvae with its gut content exposed. A blue dot was placed next to each rotifer to emphasize what they appear like under magnification and assist with identification, this larva has nine rotifers. Note forceps in corner used to maintain pressure on the slide and exposure of the gut. |
Results
Broodstock reproductive habits
Of the thirteen pairs, only eight ever produced clutches of eggs, and only five of those eight spawned consistently. From these five reliable pairs, there was still variability in the mean days between spawning (Table 1) from a low ~7 days between spawns up to ~11 days. Fortunately though, these pairs were very successful parents as all of them had >85% successful clutches, meaning they did not eat the eggs prior to their expected hatch, four days post fertilization (DPF). Additionally, these clutches were very cohesive with a fairly symmetrical round shape. However, the three pairs that were inconsistently spawning had much less success with <50% successful clutches, meaning they had eaten the eggs prior to their expected hatch. The remaining five pairs never produced eggs though behavior indicative of spawning (chasing, and male present within the tube) were observed.
Given the limited time and resources, not every pair had a hatched and observed clutch but generally hatching was >70% though one clutch only had ~5% hatch. Of these hatches there was limited variability in larval quality as each was active and alert with up to 5% of the clutch exhibiting a constant circular swimming pattern. These “spinning” larvae were excluded from the feeding trials and final experiment to avoid any confounding variables within the data.
| Table 1. An accumulation of the reproductive habits of the eight pairs that successfully spawned. Spaces left blank could not be calculated due to insufficient data. | ||||||||
| Tank 1 | Tank 3 | Tank 4 | Tank 7 | Tank 8 | Tank 9 | Tank 11 | Tank 12 | |
| Total number of spawns | 8 | 10 | 7 | 9 | 2 | 13 | 2 | 1 |
| Mean days ±SE between Spawns | 8.9±0.3 | 8.6±0.9 | 11.1±1.8 | 8.6±0.7 | 76 | 7.3±0.1 | - | - |
| Percent successful clutches | 87.5% | 100.0% | 85.7% | 88.8% | 50.0% | 92.3% | 0% | 100% |
| Mean percent ± SE of observed hatches | 32±27 | 80±0 | - | 90±0 | - | 65±18 | - | - |
Broodstock reproductive behavior: recording
At approximately 3:45 the male started exhibiting behaviors different to the previous hours. The female which remained off-screen the majority of the time was near the surface, likely hiding within the seagrass provided within the tank. The male began an erratic swimming pattern; it was almost frantic in nature swimming. He would abruptly swim toward the surface (presumably toward the female) and then dive toward the PVC den. This happened three times within a minute and then he stayed within the den for another 5 minutes before he began again. This time during the dives toward the den the angle was slightly different so it was more apparent. During his dive, he was swimming in a side to side, zig-zag like pattern, waving his tail dramatically before turning back to watch for the female to follow. This entire process occurred a total of three times before the female entered the screen. It was very apparent that she had a full clutch of eggs, as her sides were very swollen. As she approached the den the male continued his frantic swimming toward her then the den. Upon arrival both fish entered the tube and did not exit for 25 minutes. During this time the spawning likely took place as eggs were confirmed later.
Larvae First Feed
Generally, rotifers were present in the gut at 3X the density of copepods (Fig. 6) regardless of prey density during feeding. However, each treatment had slightly different gut densities. In regards to copepod treatments there is a fairly obvious linear relationship as prey density increased from 0.1/ml to 0.5/ml to 1/ml, so too did the gut content (0.5±0.06 n=75, 1.2±0.1 n=76, and 1.6±0.2 n=75). Contrary to expectation, the density of rotifers in the gut seemed to peak at 20/ml (6.0±0.7 n=78) and was lower at 2/ml (4.9±0.6 n=75) and 40/ml (5.1±0.6 n=75). The first ANOVA test was intended to illuminate if these differences were significant. * The results of the analysis (Table 2) indicate that in fact there are significant differences in gut content between prey type, prey density, as well as a significant interaction of prey type and its density. It is also worth noting that the interaction of replicate and prey type was not significant. Because the replicates p-value was so close to being significant, it was investigated further by analyzing each prey type separately. This separate analysis was unable to indicate if there were differences between prey types but confirmed that the differences noted due to different prey densities was significant. Additionally, these tests also supported the conclusion that there was no significant difference between replicates of each treatment (Tables 3&4).
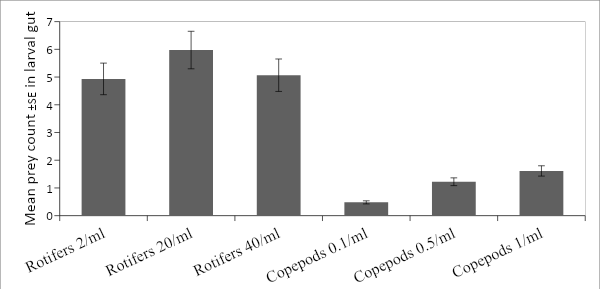
|
| Figure 6. Prey count in larval gut. |
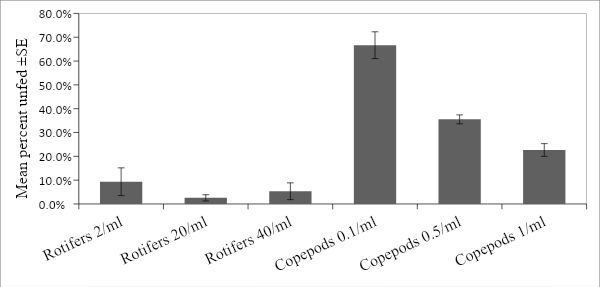
Another interesting find within the data was a varied amount of larvae within each replicate that had an empty gut and were assumed to have not eaten (unfed). Similar to gut density, this seemed to vary between prey type, and prey density (Fig 7). Specifically, copepod treatments had 2-6X more unfed larvae than rotifers; and in both prey options the percent unfed was the inverse of the gut concentration. Copepods had a negative linear relationship between percent unfed and prey density while rotifers had the lowest percent unfed at a density of 20/ml. To confirm if these observations were significant, ANOVA was used. Similar to gut concentration the percent unfed proved to be significantly different between prey, prey density, and the interaction of prey type and prey density (Table 5).
|
| Figure 7. Unfed proportion of larvae. |
*Note: Levene’s test indicated that there was not homogeneity of variance. No degree of transformation brought the data within homogeneity. I believe this is likely due to the large differences between copepods and rotifers with a particular emphasis on the amount of “unfed larvae” within the copepod treatments. To confirm the results of primary ANOVA test, the data was divided and analyzed separately in regard to copepods and rotifers. Under this regime, the rotifers passed the Levene’s test and after a log10 transformation copepods also passed.
Discussion
Broodstock reproductive habits
Beginning with the broodstock; it was apparent that despite the effort made to ensure similar conditions with feeding and water treatment, there was a significant amount of variability between breeding pairs. This is clearly documented with the differences in reproductive frequency between the spawning eight pairs as well as the complete absence of spawning in the remaining five. However, it is important to note that of the eight spawning pairs, only one was a “new pair” introduced to each other at the beginning of this experiment. This coupled with the lack of any obvious sexual dimorphism suggests that the absence of spawning may in fact be due to improper pairing of fish of the same sex. While this potentially would not be a problem for a longer study, as P. fridmani is capable of sex change in both male to female and female to male directions, for this relatively short term study the respective fish may not have completed sex conversion if they were paired with a fish of the same sex (Mies et al. 2014; Wittenrich & Munday 2005). Unfortunately, shortly after the completion of the study when Tank 12 was being cleaned, a second PVC pipe was found with eggs. This second pipe was likely spawned in throughout the experiment as a result of the disturbance surrounding the primary pipe and could be a significant contribution to that pair’s recorded poor success. This was one large drawback of the large tanks, accounting for its contents was difficult and obviously not all the potential spawning “dens” were removed prior to the beginning of the study.
Also interesting to note was the limited success of the clutches from the new pair. The clutches rarely lasted more than two days prior to being eaten. Additionally, these clutches were very loose, non-uniform in shape, and were more spaced out that the other successfully spawning pairs. This result is consistent with previous studies that showed upon pairing, there was a delay in successful clutches (Mies et al. 2014). The absence of spawning and poor clutch success in the new pairs might be a manifestation of this “learning period” during which the fish struggle to spawn successfully, or maintain a clutch until hatching. In this previous study it was noted that this learning period, though highly varied, can be quite extensive (>24 weeks).
Nonetheless, this learning period cannot account for the absence of spawning in the two pairs that had previously been established. In their case in might be that some environmental cue wasn’t quite right. Interestingly both of these pairs (Tanks 6 and 8 in Fig. 1) were likely receiving less natural direct light than the tanks (Tanks 1, 3, 4, 7 and 9). The differences in natural direct light were due to a hard plastic (opaque) cover over the tanks. This cover provides protection from rain and debris but likely modifies the amount of light received particularly to the tanks closer to the center which have less exposure from the early and late hours when the angle provides direct illumination. All of the successfully spawning pairs were near the exterior of the tank array, where natural light was the greatest and the amount of shading from the overhead cover was the least. This could suggest that light plays a role in the successful spawning, or could be random occurrence. Another potentially confounding variable was differences in temperature. While sampling to determine the presence and absence of eggs it was noted that the temperature within individual tanks often varied. Swings in temperature were likely caused by variation in water flow which could vary as a result of maintenance or algal growth. Regardless, to avoid further contaminating the results the water flow rates were left constant throughout the experiment. Further studies specifically targeting the effects of water temperature and light intensity and its effect on the reproductive habits of P. fridmani could be enlightening; given the qualitative observations collected during this study and previous studies showing fish can have varied reproductive activity under different temperature and light regimes (Chapman et al. 2009; Hovel et al. 2016). An additional cause of variation in reproduction could be resulting from the different tank sizes (120L v 2000L). Without a more balanced sample to specifically measure this, there is no real conclusion that could be made about how it might be influencing their reproductive behavior.
Another variable of interest that was left un-sampled due to limitations of the study was the “spinning” behavior noticed within the various clutches that were hatched. This behavior was found predominantly in one pair and was fairly consistent in its abundance, approximately 5-10% of the larvae. This could likely be a developmental issue or resulting from nutritional deficiencies but without further investigation it is difficult to say. Furthermore, these spinning larvae were intentionally excluded from this experiment so it is unclear whether they would be able to feed and develop naturally or if they would be incapable and die. Concerning the very poor instance of hatching at 5%, it is hypothesized that this poor hatch was result of checking the eggs to early. Despite leaving the rest of the egg bundle to hatch overnight, after the lid was removed and the larvae and egg bundle were reexposed to light there were no additional hatchings. This highlights the sensitivity of the eggs to light and indicates caution needs to be taken during this process to ensure the hatching is as successful as possible. Other studies have also noted that light intensity plays a large role within the hatching, and that hatching could be induced with higher than normal intensity light as opposed to darkness (Mies et al. 2014).
Despite the limited quantitative conclusions available from the P. fridmani broodstock study, it is clear that this species would likely be commercially viable once successful breeding pairs are established. The frequent spawning of up to twice a fortnight would allow for large production capability. This paired with fairly high hatching rates > 80% with clutch sizes in excess of 1,000 provides a solid foundation for commercial availability. Qualitatively, the data suggest that light could play an important role in successful spawning so while an effort should be made to limit photo stress, natural lighting may help P. fridmani successfully spawn regularly. However, emphasis should also be placed on regular feeding as previous studies have shown that P. fridmani is prone to eating the egg clutch prior to hatching and ensuring that each pair is well fed may reduce this occurrence (Mies et al. 2014). Finally, it would be most beneficial to have the pairs established immediately as new pairs can take a considerable amount of time prior to successfully spawning, and this could easily be exacerbated by pairing fish of the same sex. Additionally, these new pairs appear to have a hard time caring for a clutch until hatching and because of this hatching quality wasn’t even available for the new pair within this study.
Larvae First-Feed
The first-feeding experiment provided much more quantitative information than the broodstock observation. Starting with the broad results; it was clear when comparing the two prey items that rotifers were eaten in higher numbers than copepods. This is likely a consequence of a number of things. The first being the differences in prey size, as the larvae appeared incapable of ingesting more than three copepods while up to ten rotifers could be ingested comfortably. The second being prey availability within the treatments as at the highest density copepods were still half as dense as the lowest rotifer density. Lastly being the evolutionarily natural food item of P. fridmani larvae, which is more likely to be copepods as their nutritional profile and swimming behavior make them a much better prey item for larvae (Vu et al. 2017). Given the stark difference in availability of rotifers, and the lack of a mixed culture to determine preferential feeding, the fact that rotifers are consumed in high numbers is rather unsurprising and not incredibly informative. However, within each prey item the differences between treatment densities were significant and more informative.
The larval feeding response to rotifers at different concentrations was arguably the most interesting result of the study. As expected the larval gut content was lower (4.9±0.6), at the lowest feeding density of 2/ml. When density was increased to 20/ml there was a subsequent increase in the mean gut content (6.0±0.7). In following that pattern, it was hypothesized that mean gut content would continue to increase. However, at the highest concentration (40/ml), there was a drop in the mean gut content (5.1±0.6). This suggests that with increasing prey concentration, the feeding ability of P. fridmani is able to keep pace up to a certain point (between 20-40/ml). Within this range, the rotifer density appears to inhibit feeding, causing the observed decrease in gut content when compared to the 20/ml density. It might be this decrease is due to rapidly diminishing water quality at such a high density. Another possibility could be that with the increased density the amount of light is reduced to the point prey identification becomes more difficult, or the overabundance of prey might inhibit P. fridmani’s ability to identify and single out a target properly. While this study is unable to distinguish or inform what parameter is causing the reduced feeding in the 40/ml treatment, further investigation likely could by using a flow-through setup to limit any changes in water quality. Another option could be manipulating the green water with the understanding that the rotifer density could be influencing the light intensity within the smaller research vessels. Finally, it is interesting to note that previous experiments have rarely used feeding densities this high, so information on the cause and consistency of results across species is limited (Degidio et al. 2017; Hilder et al. 2014; Hilder et al. 2016). Despite the statistical significance of these results, the practical applications would likely be limited. To determine the commercial viability of these separate feeding densities a longer trial establishing survival and growth rates would be needed.
The copepod treatments were more similar to what might have been expected. As prey density increased, the average gut content of the larvae also increased. This pattern held even at the highest density which also had the highest mean gut content. This suggests that the feeding capability of the larvae had not yet been exceeded and with further increases in prey density it might be expected that the gut content might continue to rise or plateau given that the gut seemed near max at three copepods. These results are consistent with other first feed studies (Degidio et al. 2017; Hilder et al. 2014; Hilder et al. 2016). Unfortunately, given that copepods are more difficult to culture commercially in large numbers, providing copepods at densities high enough to bring the unfed percent below 10% might be impractical (Vu et al. 2017). For reason, this experiment essentially shows that if co-feeding is the most realistic option. Due to complications with prey identification post ingestion, this experiment did not include a mixed prey treatment to determine if P. fridmani showed feeding preferences for copepods vs rotifers and therefore could not determine the relative feeding efficiency of mixed culture feeding. These would be helpful studies for the future as they could inform how differences in prey density might affect feeding within mixed cultures. Given that the pilot trial showed identifying prey post ingestion can be difficult, it is recommended that dyes be used during prey culturing to allow simple and precise identification during microscopic analysis.
Another intriguing, and this time unexpected, result of the experiment was the amount of larvae within each treatment that had not eaten at all. Comparing the mean percent of unfed larvae within each treatment revealed an interesting pattern. As the mean gut content increased the percent of unfed fish decreased; more simply put, the percent unfed was the inverse reciprocal of the mean gut content. This pattern was consistent with both copepods and rotifers. At first glance this might appear odd, but intrinsically it makes sense. As the mean gut content of the larvae increases (a proxy for feeding efficiency) it is expected that the number of fish unable to successfully feed would decrease. Ultimately though, this is an easier way to see if the density provided to the larvae is enough to ensure high survival. With this study specifically, the percent unfed after four hours within the optimal rotifer density (20/ml) was less than 3% suggesting that the larvae will likely have no problem continuing to grow. If this is compared to the 22% percent unfed of the highest copepod treatment (1/ml) the need for co-feeding becomes very obvious. Using the percent unfed would be particularly useful if testing a mixed culture feeding, as the different sizes of copepods and rotifers prevents a good comparison of individuals/gut.
Conclusion
Ultimately, this study is another stepping stone in the journey to understanding the optimal culturing conditions of P. fridmani and similar marine ornamentals. This study showed that with careful attention to broodstock conditions, and a little luck, (no same sex pairs) the commercial viability of this fish can likely be established within 24 weeks. Once founded, successful spawning pairs might be expected to produce clutches of eggs every 7-11 days with an 85% chance of the clutch maturing to hatch without being eaten. Once a stable larval production has been established the ideal feed concentration for rotifers for their first feed was determined to be near 20/ml. Caution should be used above 20/ml due to the risk of feeding inhibition. If copepods are available, they were shown to be likely impractical as a solo feed option, but are otherwise well suited to cofeeding since copepods are more nutritious. Finally, future research topics concerning the production of P. fridmani were discovered, including broodstock conditions, survival and growth rate, and cofeeding of different prey items. Of these, the most important to commercial viability likely surround the broodstock. The availability of P. fridmani to market has the highest potential of improvement if light and temperature can be manipulated in such a way as to reduce the time needed for a pair to become established.
Acknowledgments
I would like to take this opportunity to extend a thank you to Chaoshu Zeng who provided this research topic and without his aide none of this would have been possible. I would also like to thank Bill Chen, Rachel Johns, and Sam Kentwell who all helped to monitor the presence and absence of eggs during the broodstock observation and occasionally went the extra mile and helped with an experiment. They played a vital role particularly when life got in the way and I needed an extra hand. Bill was particularly helpful in running trials and experiments and I hope the progress I’ve made will help contribute to his PhD.
APPENDIX
| Table 2. ANOVA summary statistics of prey and density. | |||||
| Source | Sum of Squares | df | Mean Square | F | P |
| Prey | 2020.61 | 1 | 2020.61 | 496.395 | 0.00* |
| Density | 62.04 | 2 | 31.02 | 7.62 | 0.00* |
| Replicate(Density) | 51.98 | 6 | 8.66 | 2.13 | 0.05 |
| Prey*Density | 34.70 | 2 | 17.35 | 4.26 | 0.02* |
| Prey*Replicate(Density) | 33.08 | 6 | 5.514 | 1.36 | 0.23 |
| Error | 1774.77 | 436 | 4.07 |
| Table 3. ANOVA summary statistics of log10 transformed copepod treatments | |||||
| Source | Sum of Squares | df | Mean Square | F | P |
| Density | 2.20 | 2 | 1.10 | 22.66 | 0.00* |
| Replicate(Density) | 0.19 | 6 | 0.03 | 0.65 | 0.69 |
| Error | 10.55 | 217 | 0.05 |
| Table 4. ANOVA summary statistics of rotifer treatments | |||||
| Source | Sum of Squares | df | Mean Square | F | P |
| Density | 47.15 | 2 | 23.57 | 3.38 | 0.04* |
| Replicate(Density) | 79.93 | 6 | 13.32 | 1.91 | 0.08 |
| Error | 1529.35 | 219 | 6.98 |
| Table 5. ANOVA summary statistics of proportion unfed larvae | |||||
| Source | Sum of Squares | df | Mean Square | F | P |
| Prey | 5801.58 | 1 | 5801.58 | 128.4 | 0.00* |
| Density | 1918.94 | 2 | 959.47 | 21.24 | 0.00* |
| Prey*Density | 1231.51 | 2 | 615.75 | 13.63 | 0.00* |
| Error | 542.21 | 12 | 45.18 |
References
- Abraham, S., Dinesh, R., Saseendran, S., 2017. Amphiprion Species - A key for a sustainable marine ornamental fish trade. Journal of Aquaculture in the Tropics 32, 181–188.
- Chapman, B.B., Morrell, L.J., Krause, J., 2009. Plasticity in male courtship behaviour as a function of light intensity in guppies. Behavioral Ecology and Sociobiology 63, 1757–1763.
- Degidio, J.-M.L.A., Yanong, R.P.E., Ohs, C.L., Watson, C.A., Cassiano, E.J., Barden, K., 2017. First feeding parameters of the milletseed butterflyfish Chaetodon miliaris. Aquaculture Research 49, 1087–1094.
- Hagiwara, A., Gallardo, W., Assavaaree, M., Kotani, T., Araujo, A.D., 2001. Live food production in Japan: recent progress and future aspects. Aquaculture 200, 111–127. –
- Hilder, P.I., Cobcroft, J.M., Battaglene, S.C., 2014. The first-feeding response of larval southern bluefin tuna,Thunnus maccoyii(Castelnau, 1872), and yellowtail kingfish,Seriola lalandi(Valenciennes, 1833), to prey density, prey size and larval density. Aquaculture Research 46, 2736–2751
- Hilder, P.E., Cobcroft, J.M., Battaglene, S.C., 2016. Factors affecting the feeding response of larval southern bluefin tuna, Thunnus maccoyii (Castelnau, 1872). Aquaculture Research 48, 2752–2766.
- Hill, J.E., Kilgore, K.H., Pouder, D.B., Powell, J.F.F., Watson, C.A., Yanong, R.P.E., 2009. Survey of Ovaprim Use as a Spawning Aid in Ornamental Fishes in the United States as Administered through the University of Florida Tropical Aquaculture Laboratory. North American Journal of Aquaculture 71, 206–209.
- Hovel, R.A., Carlson, S.M., Quinn, T.P., 2016. Climate change alters the reproductive phenology and investment of a lacustrine fish, the three-spine stickleback. Global Change Biology 23, 2308–2320.
- Mies, M., Guth, A.Z., Scozzafave, M.S., Sumida, P.Y.G., 2014. Spawning behaviour and activity in seven species of ornamental dottybacks. Journal of Zoo and Aquarium Research 2, 117–122. –
- Moorhead, J.A., 2015. Research-scale tank designs for the larval culture of marine ornamental species, with emphasis on fish. Aquacultural Engineering 64, 32–41.
- Moorhead, J.A., Zeng, C., 2011. Breeding of the forktail blenny Meiacanthus atrodorsalis: Broodstock management and larval rearing. Aquaculture 318, 248–252.-
- Morcom, S.M., Yang, D., Pomeroy, R.S., Anderson, P.A., 2018. Marine ornamental aquaculture in the Northeast U.S.: The state of the industry. Aquaculture Economics & Management 22, 49–71.
- Olivotto, I., Planas, M., Simões, N., Holt, G.J., Avella, M.A., Calado, R., 2011. Advances in Breeding and Rearing Marine Ornamentals. Journal of the World Aquaculture Society 42, 135–166.
- Ostrowski, A.C., Laidley, C.W., 2001. Application of marine foodfish techniques in marine ornamental aquaculture: Reproduction and larval first feeding. Aquarium Sciences and Conservation 3, 191–204.
- Stevens, C.H., Croft, D.P., Paull, G.C., Tyler, C.R., 2017. Stress and welfare in ornamental fishes: what can be learned from aquaculture? Journal of Fish Biology 91, 409–428.
- Sorgeloos, P., Dhert, P., Candreva, P., 2001. Use of the brine shrimp, Artemia spp., in marine fish larviculture. Aquaculture 200, 147–159.
- Vu, M.T.T., Hansen, B.W., Kiørboe, T., 2017. The constraints of high density production of the calanoid copepod Acartia tonsa Dana. Journal of Plankton Research 39, 1028–1039.
- Wittenrich, M.L., Munday, P.L., 2005. Bi-Directional Sex Change in Coral Reef Fishes from the Family Pseudochromidae: An Experimental Evaluation. Zoological Science 22, 797–803.












